| Cat. # | Size | Price | Inventory |
|---|---|---|---|
| 5392T | 20 µl | ||
| 5392S | 100 µl |
| REACTIVITY | H M R Mk |
| SENSITIVITY | Endogenous |
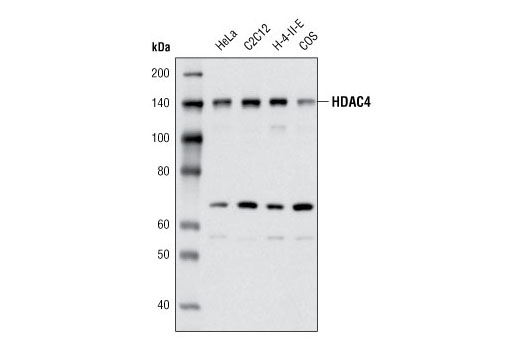
| MW (kDa) | 140 |
| Source/Isotype | Mouse IgG2a |
Product Information
| Application | Dilution |
|---|---|
| Western Blotting | 1:1000 |
| Immunoprecipitation | 1:200 |
For western blots, incubate membrane with diluted primary antibody in 5% w/v nonfat dry milk, 1X TBS, 0.1% Tween® 20 at 4°C with gentle shaking, overnight.
NOTE: Please refer to primary antibody product webpage for recommended antibody dilution.
From sample preparation to detection, the reagents you need for your Western Blot are now in one convenient kit: #12957 Western Blotting Application Solutions Kit
NOTE: Prepare solutions with reverse osmosis deionized (RODI) or equivalent grade water.
Load 20 µl onto SDS-PAGE gel (10 cm x 10 cm).
NOTE: Loading of prestained molecular weight markers (#59329, 10 µl/lane) to verify electrotransfer and biotinylated protein ladder (#7727, 10 µl/lane) to determine molecular weights are recommended.
NOTE: Volumes are for 10 cm x 10 cm (100 cm2) of membrane; for different sized membranes, adjust volumes accordingly.
* Avoid repeated exposure to skin.
posted June 2005
revised June 2020
Protocol Id: 19
This protocol is intended for immunoprecipitation of native proteins for analysis by western immunoblot or kinase activity utilizing Protein G magnetic separation.
NOTE: Prepare solutions with reverse osmosis deionized (RODI) or equivalent grade water.
10X Cell Lysis Buffer: (#9803) To prepare 10 ml of 1X cell lysis buffer, add 1 ml cell lysis buffer to 9 ml dH2O, mix.
NOTE: Add 1 mM PMSF (#8553) immediately prior to use.
A cell lysate pre-clearing step is highly recommended to reduce non-specific protein binding to the Protein G Magnetic beads. Pre-clear enough lysate for test samples and isotype controls.
IMPORTANT: Pre-wash #70024 magnetic beads just prior to use:
Transfer 20 μl of bead slurry to a clean tube. Place the tube in a magnetic separation rack for 10-15 seconds.
Carefully remove the buffer once the solution is clear. Add 500 μl of 1X cell lysis buffer to the magnetic bead pellet, briefly vortex to wash the beads. Place tube back in magnetic separation rack. Remove buffer once solution is clear. Repeat washing step once more.
IMPORTANT: The optimal lysate concentration will depend on the expression level of the protein of interest. A starting concentration between 250 μg/ml-1.0 mg/ml is recommended.
IMPORTANT: Appropriate isotype controls are highly recommended in order to show specific binding in your primary antibody immunoprecipitation. Use Normal Rabbit IgG #2729 for rabbit polyclonal primary antibodies, Rabbit (DA1E) mAb IgG XP® Isotype Control #3900 for rabbit monoclonal primary antibodies, Mouse (G3A1) mAb IgG1 Isotype Control #5415 for mouse monoclonal IgG1 primary antibodies, Mouse (E5Y6Q) mAb IgG2a Isotype Control #61656 for mouse monoclonal IgG2a primary antibodies, Mouse (E7Q5L) mAb IgG2b Isotype Control #53484 for mouse monoclonal IgG2b primary antibodies, and Mouse (E1D5H) mAb IgG3 Isotype Control #37988 for mouse monoclonal IgG3 primary antibodies. Isotype controls should be concentration matched and run alongside the primary antibody samples.
Proceed to one of the following specific set of steps.
NOTE: When using primary antibodies produced in rabbit to detect proteins with a molecular weight in the range of 50 kDa, we recommend using Mouse Anti-Rabbit IgG (Light-Chain Specific) (D4W3E) mAb (#45262) or Mouse Anti-Rabbit IgG (Conformation Specific) (L27A9) mAb (#3678) (or HRP conjugate #5127) as a secondary antibody to minimize interference produced by denatured rabbit heavy chain. For proteins with a molecular weight in the range of 25 kDa, Mouse Anti-Rabbit IgG (Conformation Specific) (L27A9) mAb (#3678) (or HRP conjugate #5127) is recommended to minimize interference produced by denatured mouse light chain.
When using primary antibodies produced in mouse to detect proteins with a molecular weight in the range of 50 kDa, we recommend using Rabbit Anti-Mouse IgG (Light Chain Specific) (D3V2A) mAb (HRP Conjugate) (#58802) as a secondary antibody to minimize interference produced by denatured mouse heavy chain.
posted December 2008
revised October 2021
Protocol Id: 121
Human, Mouse, Rat, Monkey
Monoclonal antibody is produced by immunizing animals with a recombinant protein corresponding to the amino terminus of human HDAC4 protein. The epitope corresponds to a region surrounding Gln115 of human HDAC4.
Acetylation of the histone tail causes chromatin to adopt an "open" conformation, allowing increased accessibility of transcription factors to DNA. The identification of histone acetyltransferases (HATs) and their large multiprotein complexes has yielded important insights into how these enzymes regulate transcription (1,2). HAT complexes interact with sequence-specific activator proteins to target specific genes. In addition to histones, HATs can acetylate nonhistone proteins, suggesting multiple roles for these enzymes (3). In contrast, histone deacetylation promotes a "closed" chromatin conformation and typically leads to repression of gene activity (4). Mammalian histone deacetylases can be divided into three classes on the basis of their similarity to various yeast deacetylases (5). Class I proteins (HDACs 1, 2, 3, and 8) are related to the yeast Rpd3-like proteins, those in class II (HDACs 4, 5, 6, 7, 9, and 10) are related to yeast Hda1-like proteins, and class III proteins are related to the yeast protein Sir2. Inhibitors of HDAC activity are now being explored as potential therapeutic cancer agents (6,7).
Histone deacetylases (HDACs) interact with an increasing number of transcription factors, including myocyte enhancer factor 2 (MEF2), to negatively regulate gene expression. HDACs are regulated in part by shuttling between the nucleus and cytoplasm, where export to the cytoplasm facilitates gene activation by removing HDACs from their target genes (8,9). The cytoplasmic export is facilitated by 14-3-3 proteins, which bind to specific phosphoserine residues on the HDAC proteins (8,9). These phosphoserine 14-3-3 binding modules are highly conserved between HDAC proteins, allowing for their collective regulation in response to specific cell stimuli. For example, the highly conserved HDAC4 Ser246, HDAC5 Ser259 and HDAC7 Ser155 residues are all phosphorylated by CAMK and PKD kinases in response to multiple cell stimuli, including VEGF-induced angiogenesis in endothelial cells, B cell and T cell activation, and differentiation of myoblasts into muscle fiber (10-14).
Explore pathways related to this product.
STRING - Known and Predicted Protein-Protein Interactions.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.