| Cat. # | Size | Price | Inventory |
|---|---|---|---|
| 14934S | 100 µl |
| REACTIVITY | H M R Mk |
| SENSITIVITY | Endogenous |
| MW (kDa) | 250 |
| SOURCE | Rabbit |
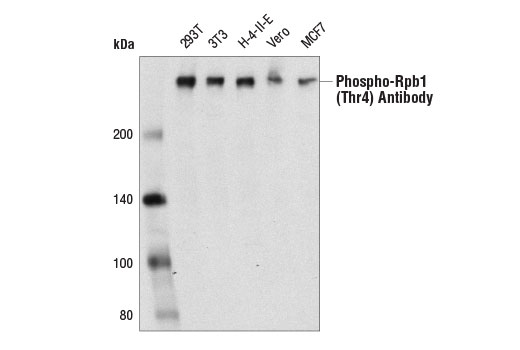
Product Information
| Application | Dilution |
|---|---|
| Western Blotting | 1:1000 |
For western blots, incubate membrane with diluted primary antibody in 5% w/v BSA, 1X TBS, 0.1% Tween® 20 at 4°C with gentle shaking, overnight.
NOTE: Please refer to primary antibody product webpage for recommended antibody dilution.
From sample preparation to detection, the reagents you need for your Western Blot are now in one convenient kit: #12957 Western Blotting Application Solutions Kit
NOTE: Prepare solutions with reverse osmosis deionized (RODI) or equivalent grade water.
Load 20 µl onto SDS-PAGE gel (10 cm x 10 cm).
NOTE: Loading of prestained molecular weight markers (#59329, 10 µl/lane) to verify electrotransfer and biotinylated protein ladder (#7727, 10 µl/lane) to determine molecular weights are recommended.
NOTE: Volumes are for 10 cm x 10 cm (100 cm2) of membrane; for different sized membranes, adjust volumes accordingly.
* Avoid repeated exposure to skin.
posted June 2005
revised June 2020
Protocol Id: 10
Human, Mouse, Rat, Monkey
Polyclonal antibody is produced by immunizing animals with a synthetic phosphopeptide corresponding to residues surrounding Thr4 of human Rpb1 CTD heptapeptide repeat. Antibodies are purified by protein A and peptide affinity chromatography.
RNA polymerase II (RNAPII) is a large multi-protein complex that functions as a DNA-dependent RNA polymerase, catalyzing the transcription of DNA into RNA using the four ribonucleoside triphosphates as substrates (1). The largest subunit, RNAPII subunit B1 (Rpb1), also known as RNAPII subunit A (POLR2A), contains a unique heptapeptide sequence (Tyr1,Ser2,Pro3,Thr4,Ser5,Pro6,Ser7), which is repeated up to 52 times in the carboxy-terminal domain (CTD) of the protein (1). This CTD heptapeptide repeat is subject to multiple post-translational modifications, which dictate the functional state of the polymerase complex. Phosphorylation of the CTD during the active transcription cycle integrates transcription with chromatin remodeling and nascent RNA processing by regulating the recruitment of chromatin modifying enzymes and RNA processing proteins to the transcribed gene (1). During transcription initiation, RNAPII contains a hypophosphorylated CTD and is recruited to gene promoters through interactions with DNA-bound transcription factors and the Mediator complex (1). The escape of RNAPII from gene promoters requires phosphorylation at Ser5 by CDK7, the catalytic subunit of transcription factor IIH (TFIIH) (2). Phosphorylation at Ser5 mediates the recruitment of RNA capping enzymes, in addition to histone H3 Lys4 methyltransferases, which function to regulate transcription initiation and chromatin structure (3,4). After promoter escape, RNAPII proceeds down the gene to an intrinsic pause site, where it is halted by the negative elongation factors NELF and DSIF (5). At this point, RNAPII is unstable and frequently aborts transcription and dissociates from the gene. Productive transcription elongation requires phosphorylation at Ser2 by CDK9, the catalytic subunit of the positive transcription elongation factor P-TEFb (6). Phosphorylation at Ser2 creates a stable transcription elongation complex and facilitates recruitment of RNA splicing and polyadenylation factors, in addition to histone H3 Lys36 methyltransferases, which function to promote elongation-compatible chromatin (7,8). Ser2/Ser5-phosphorylated RNAPII then transcribes the entire length of the gene to the 3' end, where transcription is terminated. RNAPII dissociates from the DNA and is recycled to the hypophosphorylated form by various CTD phosphatases (1).
In addition to Ser2/Ser5 phosphorylation, Ser7 of the CTD heptapeptide repeat is also phosphorylated during the active transcription cycle. Phosphorylation at Ser7 is required for efficient transcription of small nuclear (sn) RNA genes (9,10). snRNA genes, which are neither spliced nor poly-adenylated, are structurally different from protein-coding genes. Instead of a poly(A) signal found in protein-coding RNAs, snRNAs contain a conserved 3'-box RNA processing element, which is recognized by the Integrator snRNA 3' end processing complex (11,12). Phosphorylation at Ser7 by CDK7 during the early stages of transcription facilitates recruitment of RPAP2, which dephosphorylates Ser5, creating a dual Ser2/Ser7 phosphorylation mark that facilitates recruitment of the Integrator complex and efficient processing of nascent snRNA transcripts (13-15).
Phosphorylation of the Rpb1 CTD heptapeptide repeat at Thr4 in is highly conserved from yeast to mammals. However, research studies using Thr4 phosphorylation-mutant Rpb1 proteins suggest different roles for this modification among species. While phosphorylation of Thr4 in yeast is not essential (16,17), Thr4 mutants in chicken and mammalian systems result in RNA processing errors and global defects in RNA elongation (18,19). Threonine 4 is directly phosphorylated by polo-kinase3 (PLK3) and cyclin dependent kinase-9 (CDK9) activity is thought to either directly or indirectly lead to the phosphorylation of this site (18,19).
Explore pathways related to this product.
STRING - Known and Predicted Protein-Protein Interactions.
Except as otherwise expressly agreed in a writing signed by a legally authorized representative of CST, the following terms apply to Products provided by CST, its affiliates or its distributors. Any Customer's terms and conditions that are in addition to, or different from, those contained herein, unless separately accepted in writing by a legally authorized representative of CST, are rejected and are of no force or effect.
Products are labeled with For Research Use Only or a similar labeling statement and have not been approved, cleared, or licensed by the FDA or other regulatory foreign or domestic entity, for any purpose. Customer shall not use any Product for any diagnostic or therapeutic purpose, or otherwise in any manner that conflicts with its labeling statement. Products sold or licensed by CST are provided for Customer as the end-user and solely for research and development uses. Any use of Product for diagnostic, prophylactic or therapeutic purposes, or any purchase of Product for resale (alone or as a component) or other commercial purpose, requires a separate license from CST. Customer shall (a) not sell, license, loan, donate or otherwise transfer or make available any Product to any third party, whether alone or in combination with other materials, or use the Products to manufacture any commercial products, (b) not copy, modify, reverse engineer, decompile, disassemble or otherwise attempt to discover the underlying structure or technology of the Products, or use the Products for the purpose of developing any products or services that would compete with CST products or services, (c) not alter or remove from the Products any trademarks, trade names, logos, patent or copyright notices or markings, (d) use the Products solely in accordance with CST Product Terms of Sale and any applicable documentation, and (e) comply with any license, terms of service or similar agreement with respect to any third party products or services used by Customer in connection with the Products.